Medicinos ekspertas

Naujos publikacijos
Žmogaus imunodeficito virusas (ŽIV)
Paskutinį kartą peržiūrėta: 23.04.2024

Visas „iLive“ turinys yra peržiūrėtas medicinoje arba tikrinamas, kad būtų užtikrintas kuo didesnis faktinis tikslumas.
Mes turime griežtas įsigijimo gaires ir susiejamos tik su geros reputacijos žiniasklaidos svetainėmis, akademinių tyrimų institucijomis ir, jei įmanoma, medicininiu požiūriu peržiūrimais tyrimais. Atkreipkite dėmesį, kad skliausteliuose ([1], [2] ir tt) esantys numeriai yra paspaudžiami nuorodos į šias studijas.
Jei manote, kad bet koks mūsų turinys yra netikslus, pasenęs arba kitaip abejotinas, pasirinkite jį ir paspauskite Ctrl + Enter.
Įgytas imunodeficito sindromas buvo išskirtas kaip speciali liga 1981 m. Jungtinėse Amerikos Valstijose, kai daug jaunų žmonių sunkias ligas sukėlė sveikiems žmonėms nepatikslinantys ar šiek tiek patogeniški mikroorganizmai. Pacientų imuninės būklės tyrimas parodė, kad apskritai limfocitų ir ypač T-pagalbininkų skaičius smarkiai sumažėjo. Ši būklė vadinama AIDS (anglų įgytas imunodeficito sindromas - įgytas imunodeficito sindromas arba AIDS). Infekcijos gydymo būde (lytiniu keliu, per kraujo ir kraujo produktų) nurodyta infekcinės pobūdžio ligos.
Sukėlėjas AIDS buvo atrastas 1983 m nepriklausomai prancūzas L. Montagnier, kuris jį pavadino LAV Lymphoadenopathy Associated viruso), nes ji buvo rasta pacientų limfadenopatija; ir amerikietis R. Gallo, kuris pavadino virusu HTLV-III (angl. Human T-lymphotropic virus III): anksčiau jis buvo rastas I ir II limfotropinių virusų.
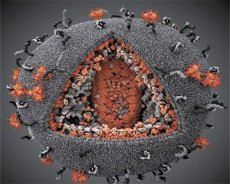
A virusų LAV ir HTVL-III savybių palyginimas atskleidė savo tapatybę, todėl, siekiant išvengti painiavos, virusas buvo 1986 metais, pavadinimas ŽIV (angl žmogaus imunodeficito viruso -. Žmogaus imunodeficito virusas arba ŽIV). ŽIV yra sferinis, jo skersmuo yra 110 nm. Virusio vokas yra daugiasluoksnės formos, sudarytos iš 12 pentagonų ir 20 šešiakampių. Centre ir kiekvienos šešiakampio yra gpl20 molekulės glikozilintas baltymų kampuose (skaičius 120 rodo, baltymų molekulinės masės į kilodaltonų). Viriono paviršiuje yra visos 72 gpl20 molekulės, kurių kiekvienas yra susijęs su intramembraniniu baltymu gp41. Šie baltymai kartu su dvigubu lipidiniu sluoksniu sudaro viriono superkapsidę (membraną).
Baltymų gpl20 ir gp41 susidaro dėl ląstelių proteazės pjovimo išankstinio baltymo Env. Proteinas gp41 formuoja stuburo "stud", kuris yra sujungtas citoplazminiu domenu su matricos baltymu p17MA tiesiai po membrana. Molekulės p17, sąveikaujančios su viriono brendimu, sudaro aštrakvą, esantį ties korpusu.
Viriono vidurinėje dalyje p24 baltymas sudaro kūginę kapsulę. Stiprinta kapsido dalis, kurioje dalyvauja pb baltymas, yra susijusi su viriono apvalkalu. Capsidoje yra dvi identiškos virusinės genominės RNR molekulės. Jie sujungiami su 5'-galais prie nukleokapsido baltymo p7NC. Šis baltymas yra įdomus tuo, kad ji turi du aminorūgščių liekanas (motyvas), cisteino-turtinga ir histidinas atomas ir kurių sudėtyje yra Zn, - jie yra vadinamos "cinko pirštas", kaip jie užfiksuoti genominės RNR molekulę, įtraukimui į virionų suformuotų. Kapsidoje taip pat yra trys fermentai. Reversas (RT) arba pol-kompleksas apima atvirkštinę transkriptazę, RNR-ase H ir nuo DNR priklausomą DNR polimerazę. Revertase yra kaip p66 / p51 heterodimeras. Proteazė (PR) - pI, prasideda ir supranta vyijų brandinimo procesą. Integracija (IN) - p31 arba endonukleazė užtikrina proviralinės DNR įtraukimą į šeimininko ląstelių genomą. Kapsidoje taip pat yra sėklinės RNR molekulės (tRNAl "3).
RNR genas ląstelėje paverčiamas atvirkštinės transkriptazės būdu į DNR genomą (DNR provirusą), kurį sudaro 9283 nukleotidų poros. Tai apribota kairiajame ir dešiniajame kampe vadinamuoju ilgo galo pakartojimu arba LTR (angl. Long long term repeat): S'-LTR - kairė ir Z'-LTR - dešinėje. LTR yra 638 nukleotidų porų.
ŽIV genomą susideda iš 9 genų, iš kurių kai kurie yra perpildyti (turi keletą skaitymo rėmų) ir turi eksonino struktūrą. Jie kontroliuoja 9 struktūrinių ir 6 reguliuojamų baltymų sintezę.
Viruso genomo LTR vertė yra ta, kad juose yra šie reguliavimo elementai, kurie kontroliuoja jo veikimą:
- transkripcijos signalas (promotoriaus regionas);
- signalas papildyti poli-A;
- Užfiksuoti signalą;
- signalo integracija;
- teigiamas reguliavimo signalas (TAT baltymo TAR);
- neigiamo reguliavimo elementas (NRE, skirtas NEF baltymui);
- sėklinės RNR (tRNA ™ 3) tvirtinimo vieta DNR minuso grandinės sintezei 3 'galuose; signalas prie LTR 5'-galo, kuris naudojamas kaip DNR pliuso grandinės sintezavimo pradmuo.
Be to, LTR yra elementai, susiję su mRNR replikacijos reguliavimu, pakuojant vRNR molekules į kapesį (Psi elementas). Galiausiai, transkribuojant genomą ilgomis mRNR, generuojami du signalai, skirti REV baltymui, kuris jungia baltymų sintezę: CAR reguliuojantiems baltymams ir KSS struktūriniams baltymams. Jei REV baltymas prisijungia prie CAR, sintezuojami struktūriniai baltymai; jei jo nėra, sintezuojami tik reguliuojami baltymai.
Viruso genomo reguliavime ypač svarbų vaidmenį atlieka šie reguliuojantys genai ir jų baltymai:
- TAT baltymas, kuris teigiamai kontroliuoja viruso dauginimąsi ir veikia per reguliuojamą TAR svetainę;
- baltymai NEV ir VPU, atliekantys neigiamą reprodukcijos kontrolę NRE svetainėje;
- proteinas REV, atliekantis teigiamą-neigiamą kontrolę. Baltymai REV kontroliuoja genų gag, pol, env ir atlieka neigiamą splicingą reguliavimą.
Taigi, ŽIV atsinaujinimas yra trigubos kontrolės - teigiamas, neigiamas ir teigiamas-neigiamas.
VIF baltymas nustato naujai sintezuojamo viruso užkrečiamumą. Jis yra susijęs su kapiliariniu baltymu p24 ir virjone yra 60 molekulių. NEF baltymas virjyje yra pavaizduotas nedideliu molekulių skaičiumi (5-10), galbūt susijęs su voku.
VPR baltymų slopina ląstelių ciklą, G2 fazės, kuriai transportavimo preintegratsionnyh kompleksų ląstelės branduolį ir aktyvuoja tam tikras virusines ir ląstelinių genus, padidina viruso replikacijos efektyvumą monocituose ir makrofagų. Biblijų VPR, TAT, REV, VPU buvimo vieta virione nėra nustatyta.
Be savo baltymų, viriono membranos sudėtis gali apimti ir kai kuriuos ląstelės-šeimininko baltymus. Baltymų VPU ir VPR dalyvauja reguliuojant virusų dauginimąsi.
Antigeniniai žmogaus imunodeficito viruso variantai (ŽIV)
Žmogaus imunodeficito virusas (ŽIV) yra labai įvairus. Net iš vieno paciento organizmo gali būti atskirti viruso štamus, kurie labai skiriasi nuo antigeninių savybių. Tokį kintamumą skatina intensyvus CD4 + ląstelių sunaikinimas ir galingas antikūnų atsakas į ŽIV infekciją. Vakarų Afrikos pacientai turi naują ŽIV formą, biologiškai arti ŽIV-1, bet imunologiškai nuo jos skiriamo, ŽIV-2. Šių virusų genomo pirminės struktūros homologija yra 42%. DNR provirusas ŽIV-2 yra 9671 bp, o jo LTR - 854 bp. Vėliau ŽIV-2 buvo izoliuota kituose pasaulio regionuose. Nėra tarpusavio imuniteto tarp ŽIV-1 ir ŽIV-2. Yra žinomos dvi didelės ŽIV-1 formos: O (Outlier) ir M (Major), pastarieji yra suskirstyti į 10 potipių (AJ). Rusijoje platinami 8 potipiai (AH).
 [
[ŽIV sąveika su ląstele
Įkvėpus kūną, virusas pirmą kartą išjuokia ląsteles, turinčias specifinį CD4 receptorių. Šiame receptoriuje yra daugybė T-pagalbininkų, mažiau - makrofagų ir monocitų, ypač virusų jautrių T-pagalbininkų.
Žmogaus imunodeficito virusas (ŽIV) atpažįsta CD4 receptorius su savo gpl20 baltymu. ŽIV sąveika su ląstelėmis vyksta pagal sekančią schemą: receptorių tarpinė adsorbcija -> ribotos fovea -> riboja vezikulą -> lizosomą. Jame viriono membrana susilieja su lizosomine membrana, o iš superkapsido išskiriamas nukleokapsidas patenka į citoplazmą; kelyje į branduolį jis sunaikinamas, o genomo RNR ir su jais susiję pagrindiniai komponentai yra išleisti. Be to, atvirkštinės transkriptazės sintezuoja dėl viriono RNR-atėmus DNR grandinės, po ribonukleaze H sunaikina viriono RNR ir virusinės DNR polimerazė sintezuoja DNR plius grandinę. DNR proviruso galuose susidaro 5'-LTR ir 3'-LTR. DNR provirusas tam tikru metu gali būti neaktyvioje formoje branduolyje, bet anksčiau ar vėliau jis integruojasi į tikslinės ląstelės chromosomą. Jame provirusas yra neaktyvioje būsenoje, kol šis T limfocitas yra aktyvuojamas mikrobinių antigenų ar kitų imunokompetentinių ląstelių. Korinio DNR transkripcijos aktyvacija yra reguliuojama specialiu branduoliniu faktoriumi (NF-kB). Tai yra DNR rišantis baltymas ir gaminamas dideliais kiekiais aktyvuojant ir proliferuojant T limfocitus ir monocitus. Šis baltymas prisijungia prie specifinių ląstelinės DNR ir panašių LTR DNR provirusų sekų bei sukelia tiek ląstelinės DNR, tiek DNR proviruso transkripciją. Sukeldamas DNR proviruso transkripciją, jis perkelia virusą iš neaktyvios būsenos į aktyvią ir, atitinkamai, patvarią infekciją, į produktyvią. Išlikti provirui neaktyvioje būsenoje gali trukti labai ilgai. Viruso suaktyvinimas yra kritinis momentas, kai jis sąveikauja su ląstele.
Nuo tos dienos, kai virusas patenka į ląstelę, prasideda ŽIV infekcijos periodas , virusas, kuris gali trukti 10 ar daugiau metų; ir nuo viruso aktyvacijos prasideda liga - AIDS. Pagal savo reguliavimo genus ir jų produktus virusas aktyviai plečiasi. TAT-baltymas gali padidinti viruso dauginimosi greitį 1000 kartų. Viruso transkripcija yra sudėtinga. Tai apima ir viso ilgio, ir subgenominių mRNR susidarymą, mRNR splaisavimą ir tolesnę struktūrinių ir reguliuojamų baltymų sintezę.
Sintezė struktūrinių baltymų įvyksta taip. Iš pradžių susintetintas poliproteino pirmtakas Pr55Gag (baltymas su M. M. 55 kD). Ji yra keturias pagrindines domenus: a matricos (MA), kapsidės (CA), nukleokapsidės (NC) ir domenų PE, iš kurių sukelti pjovimo Pr55Gag virusinė proteazės (IT samovyrezaetsya iš kitos baltymų pirmtakas - Gag-Pol) yra suformuotos atitinkamai struktūriniai baltymai p17 , p24, p7 ir pb. Poliproteino Pr55Gag susidarymas yra pagrindinė viruso dalelių susidarymo sąlyga. Būtent šis baltymas nustato viriono morfogenezės programą. Ji apima žingsnius paeiliui GAG poliproteino transporto į plazminės membranos ir jos sąveiką su baltymų-baltymų sąveikos viruso daleles ir jos jaunieji formavimas. Pr55Gag sintezuojamas laisvuose poliribomosuose; Baltymų molekulės yra transportuojamas į membraną, kuri yra įtvirtinta jų hidrofobinių regionuose. Pagrindinis vaidmuo kuriant vietinę Gag baltymo konformaciją yra CA domeno. NC-domeno jungiklis suteikia (būdu nuo "cinko pirštų") 2 molekulės genominės RNR į virusinė dalelė formavimo. Poliprodukto molekulė pirmiausia yra dimerizuota dėl matricų sričių sąveikos. Tada dimerų sujungti į hexameric (6 vienetai) kompleksų pagal sąveikos domenų CA ir NC. Pagaliau, heksamerus, jungiamieji paviršiai sudaro nesubrendusios šalutinis virionų sferinės formos, viduje, kurių sudėtyje yra virusinį genomo RNR užfiksuotą NC-domeną.
Kitą proteino prekursoriaus Prl60Gag-Pol (proteino, su m. M. 160 kDa) yra sintetinamas kaip perkeliant skaitymo rėmelis ribosomos metu vertimo Z'-galo gag geno į zoną prieš kodavimo regionas RB baltymą rezultatas. Tai Gag-Pol poliproteino apima dalinę seką Gag-baltymo (1 - 423 amino rūgščių) ir Polio sekų, kurios apima domenai PR, RT ir. Poliproteinas Gag-Pol molekulės taip pat sintezuojamos laisvose poliribomose ir transportuojamos į plazmos membraną. Poliproteinas Prl60Gagpol turi visas tarpmolekulinės sąveikos vietas, būdingas poliproteinas Gag ir membranos susiejimo vietoms. Todėl, atsižvelgiama į poliproteiną Gag-Pol saugiklio su membrana molekulės ir, kartu su Gag-molekulės apima formavimo virionų, kurie gali sukelti aktyvus proteazės ir viriono brandinimo procesas prasideda. ŽIV-1 proteazės labai aktyvi tik kaip dimeras, todėl jį samovyrezaniya iš Prl60Gag-Pol reikalauja dimerizaciją šių molekulių. Viriono brandinimas yra tas, kad išlaisvintos aktyviosios proteazės gabalai prl60Gag-Pol ir Gag55 į atpažįstamas vietas; susidaro proteinai p17, p24, p7, p6, revertase, integrazė ir jų susiejimas viruso struktūroje.
Env baltymas yra sintetinami ribosomų, susijusių su endoplazminiame retikuliaraus membranų, glikozilintas ir tada jis yra sumažinti korinio proteazę gp120 ir gp41 ir transportuojamas į ląstelės paviršiaus. Tokiu atveju gp41 perneša membraną ir prisijungia prie Gag baltymo molekulės matricos sričių, susietų su vidiniu membranos paviršiumi. Šie santykiai išlieka brandžiame virjone.
Tokiu būdu, surinkimas viruso dalelių yra agregacija pirmtako baltymų ir susijusių RNR molekulėmis, esančiomis ant membranos ląstelės šeimininko plazmos, formavimo nesubrendusių virionų ir jų išleidimo pagal jaunieji nuo ląstelės paviršiaus. Pradedant, virusas supa save su ląstelės membrana, kurioje yra įterpta gp41 ir gp120 molekulė. Per prasidėjimo ir galbūt po išleidimo viriono brendimo įvyksta, kuris yra atliekamas naudojant virusinės proteazė yra proteolitinis pjovimo Pr55Gag pirmtakų baltymų ir Prl60Gag-Pol virusas subrendęs baltymų ir jų ryšį su jomis, specifinių struktūrinių kompleksų. Pagrindinį vaidmenį viruso morfogenezės procesuose vaidina poliproteino pirmtakas Pr55Gag, kuris organizuoja ir surenka nesubrendusį virjį; Jo subrendimo procesą užbaigia specifinė virusinė proteazė.
Priežastys imunodeficito
Viena iš pagrindinių ŽIV infekcijos imunodeficito priežasčių yra masinė T-pagalbininkų mirtis. Tai įvyksta dėl šių įvykių. Pirma, dėl apoptozės miršta viruso užkrėstas T-helper virusas. Manoma, kad pacientams, sergantiems AIDS viruso replikacijos, apoptozės ir sumažėjimu, T-pagalbininkas ląstelių skaičius yra tarpusavyje. Antra, "T-ląstelėms kovotojoms atpažinti ir sunaikinti T-ląsteles užkrėsti virusu ar guolis savo adsorbuota gpl20 molekulę, taip pat virusuotų ir virusuotų, T-pagalbininkas ląstelių, kurios sudaro symplasts (syncytia) susidedančios iš kelių dešimčių ląstelių (dalis iš jie miršta dėl virusų dauginimo jose). Dėl to, kad daug T helperių ląstelių naikinimo įvyksta sumažėjimas membrana receptoriaus ekspresiją B limfocitų į IL-2, sutrikdyta sintezė įvairių interleukinų (augimo faktorių ir diferenciacijos B-limfocitų -. IL-4, IL-5, IL-6, ir kiti) dėl kurio pažeidžiama T-killer sistemos funkcija. Susilpnėja komplemento ir makrofagų sistemų veikla. Virusuotų makrofagai ir monocitai ilgai mirti, tačiau jie negalės pašalinti virusą iš organizmo. Galiausiai, dėl to, kad struktūrinių ir antigeninių panašumų su receptorių gpl20 kai epitelio ląsteles organizmo (įskaitant trofoblastų receptorių tarpininkaujant ŽIV perdavimo transplantacijos) yra sintetinamas antiretseptornyh antikūnus su plataus spektro veiksmų. Tokie antikūnai gali blokuoti įvairių korinio receptorius ir apsunkinti ligos autoimuninių sutrikimų kursą. ŽIV infekcijos pasekmė yra visų pagrindinių imuninės sistemos dalių pralaimėjimas. Tokie pacientai tampa neapsaugotu nuo daugybės mikroorganizmų. Tai veda prie oportunistinių infekcijų ir neoplastinių ligų vystymosi. Pacientams, sergantiems ŽIV infekcija, padidėjo bent trijų tipų vėžys: Kapošio sarkoma; karcinomos (įskaitant odos vėžį); B-ląstelių limfoma, atsiradusi dėl piktybinio B limfocitų degeneracijos. Tačiau ŽIV yra ne tik limfotsito- bet neurotropinė. Ji prasiskverbia CNS ląstelėse (astrocituose) arba pagal receptoriaus medijuojamo Endocitozė ir fagocitozei nustatyti astrocituose su virusuotų limfoblastais. Atsižvelgiant į viruso su astrocituose sąveikos Be to, susidaro symplasts palankias patogeno tarpląsteliniuose kanalų plitimo. Makrofaguose ir monocitų, virusas gali išlikti ilgą laiką, todėl jie tarnauja kaip rezervuaro ir jo platintojų kūno, kad galėtų įsiskverbti į visus organizmo audinius. Užkrėsti makrofagai vaidina pagrindinį vaidmenį ŽIV dreifo į CNS ir savo pralaimėjimo. 10% pacientų, sergančių pirminių klinikinių sindromų, susijusių su CNS ir parodyta su demencija (demencijos) forma. Tokiu būdu, žmonėms, kenčia ŽIV infekcijos yra būdingas 3 ligų grupes - oportunistinių infekcijų, navikinių ligų ir CNS.
ŽIV infekcijos epidemiologija
ŽIV infekcijos šaltinis yra tik asmuo - sergantis ar viruso nešiotojas. Žmogaus imunodeficito virusas (ŽIV) randamas kraujyje, spermoje, gimdos kaklelio skysčiuose; maitinančiose motinose - motinos piene. Infekcija vyksta lytiniu būdu per kraują ir jo vaistus, taip pat nuo motinos iki vaiko iki gimdymo, gimdymo metu ir po gimdymo. Viruso infekcijos atvejai per maistą, gėrimus ir vabzdžių įkandimus nėra žinomi.
Narkomanija prisideda prie AIDS plitimo. ŽIV infekcija kasmet didėja. Pasak PSO, nuo 1980 iki 2000 m. 58 milijonai žmonių buvo užsikrėtę ŽIV. Tik 2000 m. Pasaulyje užkrėstas 5,3 milijono žmonių, o AIDS mirė 3 milijonai žmonių. Rusijoje nuo 2004 m. Sausio 1 d. Užsiregistravo 264 000 ŽIV užsikrėtusių žmonių. Pusė ŽIV užsikrėtusių asmenų miršta per 11-12 metų nuo infekcijos. 2004 m. Pradžioje iš kiekvieno 100 000 Rusijos piliečių apie 180 užsikrėtė ŽIV infekcijos diagnozė. Prognozuojama, kad šiuo paplitimo lygiu bendras ŽIV infekuotų žmonių skaičius Rusijoje iki 2012 m. Bus 2,5-3 milijono žmonių. Kovos su ŽIV infekcija sudėtingumas priklauso nuo daugelio priežasčių: pirma, nėra veiksmingų gydymo ir specialios prevencijos metodų; Antra, inkubacinis ŽIV infekcijos laikotarpis gali būti ilgesnis nei 10 metų. Jo trukmė priklauso nuo T-limfocito ir DNR-proviruso, esančio jo chromosomos, aktyvacijos momento. Dar nėra aišku, ar kiekvienas AIDS užkrėstas virusas yra pasmerktas, ar gali būti ilgalaikis virusas be ligos (kuris atrodo mažai tikėtinas). Galiausiai yra keletas žmogaus imunodeficito virusų (ŽIV-1, ŽIV-2), kurių antigeniniai skirtumai neleidžia susidaryti kryžminio imuniteto. Beždžionių imunodeficito viruso (SIV) nustatymas parodo ŽIV kilmę. SIO genomo organizavimui yra panašus į ŽIV, tačiau žymiai skiriasi nukleotidų seka. ŽIV-2 serologiškai užima tarpinę vietą tarp ŽIV-1 ir SIV, o nukleotidų seka buvo arčiau SIV. Šiuo atžvilgiu V. M. Ždanovo nurodė, kad virusai ŽIV-1, ŽIV-2 ir SIV kilę iš bendro protėvio. Galima teigti, kad vienas iš SIV kažkaip pateko į žmogaus kūną, kur jis patyrė daug mutacijų, dėl kurių atsirado ŽIV-1, ŽIV-2 ir kitos jo formos.
ŽIV infekcijos simptomai
Žmogaus imunodeficito virusas pasižymi tam tikromis savybėmis, nuo kurių labai priklauso ligos patogenezė. Virusas turi labai didelį reprodukcijos greitį, kurį lemia jo reguliavimo elementai (5 000 virionų sintezuojamos per 5 minutes aktyvioje stadijoje). Dėl sulieto baltymo (gp41) buvimo virusas sukelia plataus masto sincitinių struktūrų susidarymą dėl infekuotų ir neinfekuotų T-padėjėjų susiliejimo, dėl kurio jų mirties masė. Didelės molekulės gpl20 molekulės laisvai cirkuliuoja kraujyje ir jungiasi prie neinfekuotų T-padėjėjų receptorių, dėl kurių jie taip pat pripažįstami ir sunaikinami T-killers. Virusas gali plisti per ląstelių kanalus iš ląstelės į ląstelę, šiuo atveju jis tampa mažiau prieinamas antikūnams.
Klinikiniai ŽIV infekcijos kriterijai
Suaugusiųjų ŽIV nustatyti, ar jie turi bent du rimtus simptomus kartu su bent viena simptomą nepilnametis į kitų žinomų priežasčių imunodeficitas nesant (vėžio, įgimta imunodeficitas, sunkus bado ir P. Pan.). Rimtus simptomus sudaro:
- svorio netekimas 10% ar daugiau;
- ilgai trunkanti ar nuolatinė karščiavimas;
- lėtinis viduriavimas.
Nedideli simptomai: nuolatinis kosulys, apibendrintas dermatitas, pasikartojantis herpes zosteras, burnos ertmės kandidozė ir ryklė, lėtinis herpes simpleksas, apibendrintas limfadenopatija. AIDS diagnozė nustatoma tik Kapoši sarkoma, kriptokokinis meningitas, pneumocistinė pneumonija. Klinikiniam ligos vaizdui įtakos turi oportunistinė infekcija.
Žmogaus imunodeficito viruso (ŽIV) kultivavimo metodai
ŽIV-1 ir ŽIV-2 gali būti kultivuojami tik vieno TCB4 limfocitų klonuose - H9, gautuose iš leukeminių TCV4 limfocitų. Tuo pačiu tikslu taip pat gali būti naudojamos vienkartinės astrocitų ląstelių kultūros, kuriose ŽIV-1 daugina gerai. Nuo gyvūnų iki ŽIV-1 jautrių šimpanzių.
Atsparumas viruso išorinei aplinkai yra nedidelis. Jis miršta saulės spindulių ir ultravioletinių spindulių poveikio metu, 30 minučių sunaikinamas 80 ° C temperatūroje, kai gydomas dažniausiai naudojamais dezinfekavimo priemonėmis - 20-30 minučių. Norėdami dezinfekuoti virusą turinčią medžiagą, būtina naudoti mikobaktericidinius dezinfekavimo priemones, nes jie yra efektyviausi mikroorganizmams su didžiausiu pasipriešinimu.
Laboratorinė ŽIV infekcijos diagnozė
Pagrindinis būdas diagnozuoti virusą ir ŽIV infekciją yra fermentinis imuninis tyrimas. Tačiau, dėl to, kad gpl20 turi struktūrinę ir antigeninį panašumo į receptorių tam tikrų žmogaus ląstelių, įskaitant receptorių, kad vykdo transportą imunoglobulinų per epitelinių ląstelių gleivinių organizme gali būti rodomi antikūnų, susijusius antikūnus prieš gpl20. Tokiu atveju gali būti klaidingai teigiami IFM rezultatai. Todėl visi teigiamai reaguojantys tiriami serumai yra papildomai analizuojami imunoblotiniu metodu arba Vakarų blotingai. Šis metodas pagrįstas antikūnų, kurie turi būti tiriami po elektroforezės atskyrimo ir vėlesnio bandymo su paženklinti antivirusiniais antikūnais, identifikavimo. Virusologinis metodas yra mažai naudingas dėl viruso kultūros sudėtingumo. H9 limfocitų klonas yra naudojamas virusinių antigenų - būtinų diagnostikos bandymų sistemų komponentų - gauti. CDR metodas leidžia aptikti virusą ankstyvoje viremijos stadijoje.
ŽIV infekcijos gydymas
Būtina rasti arba sintetina vaistai veiksmingai slopina atvirkštinės transkriptazės aktyvumą (atvirkštinės transkriptazės), arba virusinės proteazę. Jie būtų užkirstas kelias proviral DNR ir (arba) išsidėstymą slopina ląstelėje replikaciją viruso. Modernus strategija gydymo ŽIV infekuotų pacientų remiasi kombinuoto narkotikų vartojimo, kurie slopina viruso proteazės (vienas iš narkotikų) ir revergazu (2 įvairių narkotikų vartojimo) principo, - kartu (trigubos) terapija. Rusijoje, dėl ŽIV infekuotų žmonių gydymui rekomenduojama kartu naudoti du vietos vaistai: Crixivan Phosphazide ir specifiškai slopina dauginimąsi ŽIV pradžioje ir pabaigoje etapuose veisimo, ypač riboto aktyvumo AZT.
Specifinė profilaktika problemos slypi būtinybė sukurti vakciną, kuri užtikrintų formavimąsi veiksmingą lasteliniame imuniteto dėl viruso specifinis citotoksinių limfocitų pagrindu be reikšmingų antikūnų gaminimąsi. Toks imunitetas teikti Thl-pagalbininkas ląsteles. Ji yra įmanoma, kad antikūnas, įskaitant neutralizavimo, ne tik yra veiksminga slopinant ŽIV infekciją, bet ne aukšto lygio slopina ląstelių medijuotos imunitetą. Todėl, anti-ŽIV vakcinos turi atitikti pirmiausia du pagrindinius reikalavimus: a) turi būti visiškai saugūs ir b) skatinti citotoksinių T limfocitų aktyvumą. Tiriamas įvairių įgyvendinimo variantuose vakcinų gaunama iš paskerstų (inaktyvuota) viruso ir atskirų antigenų su aukšto apsauginėmis savybėmis efektyvumą. Šie antigenai arba gali būti išskirtas iš virionų patys arba cheminės sintezės būdu. Vakcina, kurią sukūrė genų inžinerijos metodais. Ji yra rekombinantinis karvių raupų virusas vykdyti ŽIV genus, atsakingas už antigenų sintezės su stiprių imunogeninių savybių. Sprendimas dėl šių vakcinų veiksmingumo reikalauja nemažai laiko, nes ilgą trukmės inkubacinis laikotarpis, ŽIV infekcijos ir didelio kintamumo patogeno. Sukūrimas labai veiksmingas ŽIV vakcinos - skubiai esminė problema.